Статтю опубліковано на с. 88-94
Респіраторна патологія — актуальна проблема медицини. У наш час все частіше зустрічаються труднощі в проведенні антибактеріальної терапії — зниження ефективності застосування антибактеріальних препаратів в клінічній практиці. Серед ряду причин такого становища є й утворення біоплівок у дихальних шляхах при респіраторних інфекціях. Біоплівка — найбільш поширена форма організації життєдіяльності мікроорганізмів, що захищає їх від несприятливих фізичних, хімічних і біологічних факторів (Nenke P. et al., 2006; Vu B. et al., 2009). Біоплівкові інфекції погано реагують на стандартну терапію антибіотиками, і їх лікування становить серйозну проблему в клінічній практиці.
Мета роботи — узагальнити дані щодо розвитку біоплівок у дихальних шляхах при респіраторній патології та впливу на них амброксолу.
У складі біоплівки частка самих бактерій, за даними різних авторів, становить від 5 до 35 % (Kostakioti M., 2013). Іншу, більшу частину займає міжклітинний матрикс, що являє собою складну біохімічну суміш полісахаридів, глікопептидів, нуклеїнових кислот і ліпідів (Flemming H.C., 2010; Tetz V.V. et al., 2004; Hall-Stoodley L., 2004; Høiby N. et al., 2011).
Матрикс виконує захисну роль, а також забезпечує бактеріям можливість обмінюватися «інформацією» між собою. Застосування засобів скануючої мікроскопії дозволило наочно побачити неоднорідну структуру різних шарів біоплівки, у якій можна виділити транспортні й водні канали, відзначити певні закономірності у розподілі різних видів мікроорганізмів (Yung-Hua Li, 2012). Крім абсолютного біологічного синергізму, коли продукти життєдіяльності одного виду слугують харчовим середовищем для іншого, у біоплівці спостерігається й чітко побудована взаємодія між мікробами, що належать до одного або різних видів, за допомогою спеціальних сигнальних систем. Завдяки наявності позаклітинних ферментів матрикс може розглядатися як зовнішня травна система, у якій також виявляється акумуляція води й харчових речовин (Kostakioti M., 2013).
Той факт, що бактерії здатні утворювати складні бактеріальні спільноти, був відомий давно, проте в 90-х роках минулого сторіччя було доведено існування феномену колективної поведінки бактерій, який отримав назву quorum sensing (QS) — «відчуття кворуму» (Jakobsen T.H. et al., 2011; Hall-Stoodley L. et al., 2009; Tian X., 2012; O’Toole G., 2000). Quorum sensing відбувається за участі регуляторних систем за рахунок взаємодії зі специфічними рецепторами, розташованими на поверхні мембрани бактерії, такими як N-ацил-гомосериндегідрогенази лактон (Schuster M., 2013) та інші (Bhardwaj A.K., 2013).
Важливою особливістю біоплівки є її гетерогенність: деякі мікроорганізми в ній перебувають у неактивному стані і є більш стійкими до дії зовнішніх факторів, інші швидко ростуть і продукують компоненти матриксу (Sauer K et al., 2002; Stewart P.S., 2008).
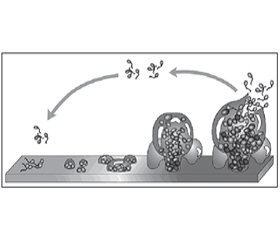
Життєвий цикл біоплівки містить декілька стадій (рис. 1) (Pintucci J.P., 2010).
Формування біоплівки починається з адгезії (прилипання) бактерій до поверхні за допомогою поверхневих адгезинів і олігосахаридів на клітинних мембранах і формування мікроколоній. Потім відбувається дозрівання біоплівки, що характеризується синтезом матриксу й формуванням складних відносин між мікроорганізмами. Тільки зріла біоплівка починає виділяти мікроорганізми за свої межі й таким чином поширюватись на нові території.
Близько 60 % хронічних і рецидивуючих інфекцій, що торпідні до стандартної терапії, пов’язані із формуванням на різних поверхнях (передусім на слизових оболонках) біоплівок (Pintucci J.P., 2010; Nazzari Е., 2015; Matthew R., 2003).
На сьогодні доведена роль мікробних біоплівок в етіології й патогенезі багатьох гострих і хронічних бактеріальних інфекцій, особливо останніх. До таких захворювань відносять інфекції сечових шляхів (Shan K.D. et al., 2004), інфекції середнього вуха (Costerton J.M. et al., 1999), муковісцидоз (Davies J.C., 2009) і вентиляційно-залежні пневмонії (Wiener-Kronish J.P., 2008), інфекційний ендокардит, інфекції протезованих клапанів, катетер-асоційовані інфекції, стоматологічні захворювання (карієс, пародонтит, гінгівіт).
Основними бактеріями, що утворюють біоплівку при хронічних захворюваннях, є S.aureus, Enterococcus, Streptococcus, Klebsiella, Pseudomonas, E.coli.
Такі бактерії, як Salmonella, E.coli, Y.entero–calitica, Listeria, Campylobacter, виявляють у складі біоплівок на поверхні харчових продуктів або ємностей для їх зберігання.
Розвиток біоплівки на поверхні медичних пристроїв — протезів, катетерів, імплантів — є етіопатогенетичною основою так званих девайс-асоційованих інфекцій. У Німеччині, де ведеться статистичний облік девайс-асоційованих інфекцій, кількість подібних захворювань перевищує 100 000 випадків за рік. У Західній Європі і США щорічно реєструється понад 500 000 випадків катетер-асоційованих інфекцій.
Підтверджено формування біоплівок у дітей із розростанням аденоїдних вегетацій, особливо у випадку розвитку на їх тлі рецидивуючих середніх отитів (Saafan M.E., 2013; Saylam G., 2010), а також при хронічних гіперпластичних тонзилітах. Так, серед 22 дітей з повторюваними загостреннями хронічного гіперпластичного тонзиліту біоплівки було знайдено в 50 % випадків, причому основним збудником був золотистий стафілокок (81,8 %) (Torretta S., 2014).
Відомо, що від 67 до 78 % клінічних ізолятів золотистого стафілококу можуть формувати біоплівки. Серед метицилін-резистентних золотистих стафілококів частка біоплівкоутворюючих штамів доходить до 96 % (Singh R. et al., 2010).
Наукові дослідження показали, що при хронічному синуситі золотистий стафілокок виділяють у 50 % випадків, гемофільну паличку — у 28 % випадків, синьогнійну паличку й гриби — у 22 % (Foreman A. et al., 2009).
Формування біоплівок за участю золотистого й епідермального стафілококу може відігравати вирішальну роль у патогенезі остеомієліту, синуситів і риносинуситів, ендокардитів, отитів, муковісцидозу, септичних артритів, хронічної ранової інфекції.
Дослідження інституту Пастера (Франція) у 2010 році вперше показали, що деякі віруси, зокрема вірус HTLV-1, здатні утворювати комплекси, схожі на бактеріальні біоплівки, тим самим захищаючи себе від дії імунної системи. Вивчення вірусних біоплівок може визначити нову терапевтичну стратегію, що буде орієнтована не тільки на вірус, але й на вірусну біоплівку.
Особливу роль відіграють біоплівки в етіології та патогенезі захворювань дихальної системи.
Розвиток біоплівок при муковісцидозі обумовлений деякими факторами. У першу чергу він пов’язаний із погіршенням мукоциліарного кліренсу (Ratjen F.A., 2009). Також певну роль відіграє підвищена щільність рецепторів на поверхні клітини, що здатні зв’язувати бактерії, і порушення фагоцитозу бактерій епітеліальними клітинами (Davies J.C., 2009). Гіпоксія та слизові пробки сприяють формуванню біоплівки, оскільки створюють несприятливі умови для планктонних форм бактерій і сприяють їх перетворенню на стійкі «сидячі» форми (Boucher R.C., 2004). При муковісцидозі найчастіше у складі біоплівок знаходять гемофільну паличку й золотистий стафілокок. Проте також зустрічаються й інші бактерії. Наприклад, у 10 % хворих на муковісцидоз у біоплівках виявляють Burkholderia cepacia (Davies J.C., 2009).
Накопичені дані свідчать про те, що біоплівки можуть відігравати також певну роль при хронічних обструктивних хворобах легень (ХОЗЛ) і бронхоектазах (Mannino D.M., 2002; Buist A.S., 2007; Lozano R., 2012).
Основними бактеріями, що входять до складу біоплівки при ХОЗЛ, є, як правило, H.influenzae, пневмокок і Moraxella catarrhalis (Sethi S., 2007). Механізм формування біоплівки при ХОЗЛ пов’язаний з пошкодженням епітелію дихальних шляхів через куріння тютюну або забруднення довкілля (Eldika N., 2006). Ці фактори сприяють розвитку запалення та активації макрофагів, нейтрофілів і дендритних клітин (Brusselle G.G., 2011; Rovina N., 2013). Останнє може також активувати клітини адаптивного імунітету й запускати автоімунні механізми, що викликають хронічне запалення (Brusselle G.G., 2011; Dancer R., 2013).
У мокротинні пацієнтів із ХОЗЛ відзначається високий рівень типових для біоплівки ферментів (Murphy T.F., 2005).
Як при ХОЗЛ, так і при бронхоектазії хронічна інфекція підтримується вірусами грипу, P.aeruginosa, М.catarrhalis, пневмококами й золотистим стафілококом (Chalmers J.D., 2013). Певну участь у формуванні біоплівки при бронхоектазах беруть також і нетуберкульозні мікобактерії (McNabe M., 2011; Mirsaeidi M., 2013).
Доведено, що біоплівки розвиваються протягом перших 48 годин від початку механічної вентиляції легень, у їх складі часто виявляють Acinetobacter baumannii і синьогнійну паличку (Lambiase A., 2009).
Вентиляційно-асоційована пневмонія є однією з найбільш поширених внутрішньолікарняних інфекцій у відділенні інтенсивної терапії, її виявляють у 8–28 % пацієнтів, які отримують механічну вентиляцію (Di Bonito M., 2012). Сумарна частота вентиляційно-асоційованих пневмоній коливається від 2 до 16 епізодів на 1000 днів штучної вентиляції легень, і пік захворюваності спостерігається між 5-м і 9-м днем механічної вентиляції (Barbier F., 2013).
Біоплівкові інфекції погано реагують на стандартну терапію антибіотиками, і їх лікування становить серйозну проблему в клінічній практиці. У складі біоплівки мікроорганізми набувають множинної резистентності до антибактеріальних препаратів і стають недоступними для клітинних і гуморальних факторів захисту (Pintucci J.P., 2010; Desrosiers M., 2007).
Висока стійкість мікроорганізмів біоплівок до антибіотикотерапії підтверджується тим, що мінімальна інгібуюча концентрація антибактеріальних препаратів може бути в сотні й тисячі разів вищою, ніж для планктонних форм бактерій (Mah T.-F.C., 2001).
Стійкість до антибактеріальної терапії в мікроорганізмів біоплівки обумовлена декількома факторами (Høiby N., 2010). Одним із них є погана дифузія антибіотиків всередину біоплівки, унаслідок чого концентрація антибіотика в біоплівці може бути меншою, ніж у тканинах або крові, і в зв’язку з цим недостатньою для ерадикації збудника. Крім того, частина бактерій всередині біоплівки перебуває в неактивному стані, і тому такі бактерії нечутливі до антибактеріальних препаратів, що впливають на метаболічну активність. Епідеміологічні дослідження доводять, що мікроорганізми в межах однієї біоплівки можуть обмінюватися між собою генетичною інформацією щодо антибіотикорезистентності (Donlan R.M. et al., 2002; Chebotar I.V. et al., 2012). А визначення чутливості до антибіотиків може не відображати чутливість всієї біоплівки, оскільки виявляється реакція на антибіотик активних форм збудників, а неактивних — залишається невідомою (Desrosiers M., 2007).
Бактерії біоплівки захищені від основних компонентів імунітету людини, зокрема від катіонних і аніонних антибактеріальних пептидів і фагоцитозу (Vuong C., 2004).
З огляду на вищенаведені факти в лікуванні хворих постає необхідність вдаватись до заходів, що спрямовані на запобігання утворенню біоплівок або їх видалення в найближчий час після утворення (Golub A.V., 2012).
На жаль, використання профілактичної антибіотикотерапії з метою запобігання утворенню біоплівок виявилося неефективним і в даний час не рекомендується (Coffin S.E., 2008; Masterton R.G., 2008; Muscedere J., 2008; Torres A., 2009). А вирішальне значення мають стратегії, спрямовані на покращання мукоциліарного кліренсу, вплив на імунне запалення, відновлення цілісності слизових оболонок, що запобігає адгезії мікроорганізмів до поверхні слизової оболонки і тим самим гальмує розвиток і поширення біоплівки.
В останні роки серед препаратів, що мають специфічну дію на біоплівки, відомий мукокінетик амброксол (Cataldi M., 2014).
Традиційно амброксол широко використовується як у терапевтичній, так і в педіатричній практиці при лікуванні гострих і хронічних респіраторних інфекцій завдяки його властивостям добре зменшувати в’язкість мокротиння й полегшувати його відходження за рахунок збільшення мукоциліарного кліренсу (Cataldi M., 2014; Varelogianni G., 2013). Діючи як муколітичний засіб і експекторант, він покращує функцію зовнішнього дихання. Також відома властивість амброксолу збільшувати вміст сурфактанту в легенях шляхом стимуляції його продукції пневмоцитами. Амброксол здатний запобігати розвитку респіраторного дистрес-синдрому в новонароджених, оскільки вільно проникає через плацентарний бар’єр і стимулює внутрішньоутробне утворення сурфактанту.
Амброксол підвищує концентрацію антибактеріальних препаратів у бронхоальвеолярному секреті й у зоні запального вогнища в легенях (Bonnetti P., 1993), має антиоксидантний (Zhao S.P., 2004; Nowak D., 1994) і протизапальний ефект (Cataldi M., 2014; Zhang B., 2000; Malerba M., 2008; Snowden J.N., 2012). Aмброксол чинить протизапальний ефект шляхом декількох механізмів, включаючи інгібування синтезу NO, дегрануляцію нейтрофілів і індукцію синтезу поверхнево-активної речовини. Цей протизапальний ефект може бути додатковим механізмом, що сприятливо впливає на перебіг біоплівкових захворювань дихальних шляхів.
Проте останнім часом було виявлено нові властивості амброксолу, що дозволяють розширити спектр його застосування в педіатричній практиці, у тому числі і як засобу, що здатний впливати на біоплівки.
Нещодавно отримані дані показують, що амброксол призводить до порушення структурної цілісності бактеріальної біоплівки (Li F., 2008). Скануюча електронна мікроскопія продемонструвала, що після 7 днів лікування амброксолом біоплівка, утворена P.aeruginosa, ставала тоншою і фрагментованою порівняно з тою, що формувалась у контрольних умовах.
Доведено вплив амброксолу на всі етапи розвитку біоплівки.
M.M. Hafez та співавт. (Hafez M.M., 2009) досліджували вплив різних муколітиків на біоплівку й дійшли висновку, що амброксол порівняно з бромгексином, хлоридом амонію й ацетатом амонію більш істотно перешкоджає бактеріальній адгезії в експерименті. Амброксол у низькій концентрації (2,5 нг/мл), як і ацетилцистеїн, може гальмувати первинне приєднання мікроорганізмів до поверхонь (Riise G.C., 2000; Hafez M.M., 2009) і, відповідно, початкові етапи утворення біоплівки. У більш високих концентраціях амброксол має здатність безпосередньо діяти на біоплівку, знижуючи, наприклад, продукцію альгінату, що є важливим фактором вірулентності й антибіотикорезистентності (Li F., 2008). Зокрема, показано, що в концентраціях 1,875 і 3,75 мг/мл амброксол призводить до дозозалежного зниження вмісту альгінату біоплівок Pseudomonas aeruginosa, отриманих у лабораторних умовах (Li F., Yu J., Yang H., 2008; Lu Q., Yu J., Bao L. et al., 2013).
Вищевказані фармакологічні ефекти амброксолу виявлені щодо деяких видів патогенних бактерій і не підтверджуються щодо вірусів (Ndour C.T., 2001).
Дослідження, проведені in vitro в дітей, показали, що амброксол має дозозалежний бактерицидний вплив на Haemophilus influenzae і здатний руйнувати біоплівки, до складу яких входить цей збудник (Gao X., 2014). Було показано, що концентрація амброксолу в середовищі 0,49 мг/мл є більш ефективною щодо впливу на біоплівки порівняно з концентрацією 0,25 мг/мл.
Існує гіпотеза про те, що амброксол може діяти шляхом контролю транскрипції генів оперона AlgU, хоча молекулярний механізм цього ефекту ще належить встановити.
Існують також певні докази того, що амброксол може впливати на quorum sensing бактерій (Lu Q., 2010). Було вивчено вплив амброксолу на структуру біоплівки в мутантів синьогнійної палички, позбавлених двох основних систем, що забезпечують quorum sensing, і у їх диких родичів. Виявлено, що ефект амброксолу був значно ослаблений у мутантних штамів.
Отже, амброксол впливає на формування бактеріальної біоплівки на різних стадіях і шляхом декількох механізмів, включаючи прямий антагонізм адгезії, інгібування синтезу альгінату та quorum sensing бактерій.
Інгібуючий ефект амброксолу поширюється й на грибкові біоплівки (Pulcrano G., 2012). Доведено, що амброксол впливає на ріст Candida albicans, що знаходиться у складі біоплівки при кандидозі слизових оболонок ротової порожнини (Rene H.D., 2014).
Аналізуючи результати сумісного застосування амброксолу й ванкоміцину в новонароджених із катетер-асоціованими інфекціями, Y. Zhang та співавт. (Zhang Y., 2015) дійшли висновку, що ефективність амброксолу проявляється в потенціюванні бактерицидного ефекту ванкоміціну на епідермальний стафілокок біоплівки як у пробірці, так і в природних умовах.
Автори використали модель кроликів, яким вводили катетери, вкриті біоплівкою, що містить епідермальний стафілокок. Тварини отримували лікування одного з чотирьох типів: гепарин, амброксол, ванкоміцин і комбінацію ванкоміцину з амброксолом протягом трьох днів. Електронна мікроскопія й бактеріальне дослідження показали, що аброксол проявляє значну ефективність щодо потенціювання бактерицидного ефекту ванкоміцину на епідермальний стафілокок біоплівки. У зв’язку з цим автори рекомендують комбінацію ванкоміцину й амброксолу для лікування новонароджених з катетер-асоційованими інфекціями.
З огляду на виявлені властивості амброксолу та здатність його гальмувати утворення й пригнічувати функціонування біоплівки є перспективним розширення показань до його застосування в клінічній практиці.
Оскільки не всі амброксоли мають однакову переносимість та ефективність, найбільш широко в практиці використовуються розчини й таблетки амброксолу німецької компанії «Берлін-Хемі/А. Менаріні» під ТМ «Флавамед» (Банадига Н.В., 2015).
Молекула Флавамеду має комплексну дію: муколітичну (розрідження густого секрету, що виділяється слизовою бронхів, без збільшення його об’єму), секретомоторну (покращення евакуації секрету завдяки відновленню мукоциліарного кліренсу) та антиадгезивну (збільшення синтезу сурфактанту, зменшення прилипання мокротиння до слизової оболонки). Місцевий анестезуючий ефект Флавамеду пояснюється його властивостями блокування натрієвих каналів. Експериментально доведено, що молекула Флавамеду зменшує вивільнення та/або концентрацію 13 медіаторів запалення (LTB4, cys-LT, IL-1, IL-2, IL-4, –IL-6, IL-8, IL-12, IL-13, IFN-γ, TNF-γ), гістаміну та оксидативних метаболітів (Beeh К.М. et al., 2008). Рідкі форми Флавамеду вирізняє те, що вони випущені у формі розчину (а не сиропу) без етанолу та цукру, мають меншу кількість домішок та ароматизаторів, прийнятні органолептичні властивості, що зумовлює низький відсоток побічних ефектів. Таблетки Флавамеду відрізняються маленьким розміром (діаметр таблеток лише 7 мм), що значно полегшує процес ковтання дітьми.
Відсутність у рідких формах Флавамеду спирту та цукру мотивувало його використання не лише при гострій чи хронічній бронхолегеневій патології, але й у дітей із бронхіальною астмою, коли після купірування нападу йому на зміну приходить малопродуктивний кашель (свідчення регресу основних компонентів бронхообструкції). У післянападному періоді дуже важливо відновити дренажну функцію бронхів, забезпечити ефективну евакуацію слизу, що запобігатиме можливості його інфікування. Водночас Флавамед добре поєднується не лише з антибактеріальною, але й з бронхолітичною терапією, не провокує бронхо–спазм на відміну від муколітика ацетилцистеїну. Клінічно доведено доцільність використання Флавамеду при сухому кашлі, наприклад при гострому бронхіті, коли є потреба трансформувати його у вологий, щоб прискорити темпи одужання (Банадига Н.В., 2015).
Отже, лікування захворювань, що супроводжуються утворенням біоплівок, — актуальна проблема клінічної практики. Наявні літературні дані переконливо свідчать про ефективність застосування амброксолу (Флавамеду) у дорослих і дітей при захворюваннях, що супроводжуються утворенням біоплівок. Гальмуючу дію на утворення й функціональну активність біоплівки амброксол чинить за рахунок як уже відомих властивостей (антиоксидантний, протизапальний ефект, здатність покращувати мукоциліарний кліренс), так і завдяки нещодавно виявленій здатності безпосередньо впливати на структуру біоплівки і її quorum sensing.
Список литературы
1. Barbier F., Andremont A., Wolff M., Bouadma L. Hospital-acquired pneumonia and ventilator-associated pneumonia: recent advances in epidemiology and management // Curr. Opin. Pulm. Med. — 2013. — 19. — 216e28.
2. Bhardwaj A.K., Vinothkumar K., Rajpara N. Bacterial quorum sensing inhibitors: attractive alternatives for control of infectious pathogens showing multiple drug resistance // Recent Pat. Antiinfect. Drug Discov. — 2013. — 8. — 68e83.
3. Bonnetti P. Ambroxol plus amoxicillin versus amoxicillin alone in various respiratory tract infections // Investig. Med. Internatio–nal. — 1993. — 20. — 99-103.
4. Boucher RC. New concepts of the pathogenesis of cystic fibrosis lung disease // Eur. Respir. J. — 2004. — 23. — 146e58.
5. Brusselle G.G., Joos G.F., Bracke K.R. New insights into the immunology of chronic obstructive pulmonary disease // Lancet. — 2011. — 378. — 1015e26.
6. Buist A.S., McBurnie M.A., Vollmer W.M., Gillespie S., Burney P., Mannino D.M. et al. International variation in the prevalence of COPD (the BOLD study): a population-based prevalence study // Lancet. — 2007. — 370. — 741e5.
7. Cataldi M., Sblendorio V., Leo A., Piazza O. Biofilm-dependent airway infections: a role for ambroxol? // Pulm. Pharmacol. Ther. — 2014 Aug. — 28(2). — 98-108.
8. Chalmers J.D., Hill A.T. Mechanisms of immune dysfunction and bacterial persistence in non-cystic fibrosis bronchiectasis // Mol. Immunol. — 2013. — 55. — 27e34.
9. Chebotar I.V. et al. Antimicrobial Resistance of Bacteria in Biofilms // Clinical Microbiology and antimicrobial chemotherapy. — 2012. — Vol. 14. — Р. 51-58.
10. Coffin S.E., Klompas M., Classen D., Arias K.M., Podgorny K., Anderson D.J. et al. Practice recommendation of Society for Healthcare Epidemiology of America/Infectious Diseases Society of America (SHEA/IDSA): strategies to prevent ventilator-associated pneumonia in acute care hospitals // Infect. Control. Hosp. Epidemiol. — 2008. — 29. — S31e40.
11. Costerton J.M. et al. Bacteril biofilms: a common cause persistens infection // Science. — 1999. — Vol. 284. — Р. 1318-1322.
12. Dancer R., Sansom D.M. Regulatory T cells and COPD // Thorax. — 2013. — 68. — 1176e8.
13. Davies J.C., Bilton D. Bugs, biofilms, and resistance in cystic fibrosis // Respir. Care. — 2009. — 54. — 628e40.
14. Desrosiers M., Bendauah Z., Barbeau J. Effectiveness of topical antibiotics on Staphylococcus aureus biofilm in vitro // Am. J. Rhinol. — 2007. — 21. — 149-53.
15. Di Bonito M., Caiazzo S., Iannazzone M., Miccichи V., De Marco G., De Robertis E. et al. Prognostic differences between VAP caused by Acinetobacter baumannii and VAP caused by other microorganisms // Transl. Med. UniSa. — 2012. — 3. — 15e21.
16. Donlan R.M. et al. Biofilms: survival mechanisms of clynically relevant microorganism // Clin. Microbiol. Rev. — 2002. — Vol. 15. — Р. 167-193.
17. Eldika N., Sethi S. Role of nontypeable Haemophilus influenzae in exacerbations and progression of chronic obstructive pulmonary disease // Curr. Opin. Pulm. Med. — 2006. — 12. — 118e24.
18. Flemming H.C., Wingender J. The biofilm matrix // Nat. Rev. Microbiol. — 2010. — 8. — 623-33.
19. Foreman A., Psaltis A.J., Tan L.W. et al. Characterization of bacterial and fungal biofilms in chronic rhinosinusitis // Am. J. Rhinol. Allergy. — 2009. — 23. — 556-61.
20. Gao X., Zhang Y., Lin Y., Li H., Xin Y., Zhang X., Xu Y., Shang X. Effect of ambroxol on biofilm of Haemophilus influenzae and bactericidal action // Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi. — 2014 May. — 28(10). — 721-3.
21. Golub A.V. Bacterial Biofilms — a New Therapeutic Target? // Clinical microbiology and antimicrobial chemotherapy. — 2012. — Vol. 14, № 1. — Р. 23-29.
22. Hafez M.M., Aboulwafa M.M., Yassien M.A., Hassouna N.A. Activity of some mucolytics against bacterial adherence to mammalian cells // Appl. Biochem. Biotechnol. — 2009. — 158. — 97e112.
23. Hall-Stoodley L. et al. Evolving concepts in biofilm infections // Cell Microbiol. — 2009. — Vol. 11. — Р. 1034-1043.
24. Hall-Stoodley L., Costerton J.W., Stoodley P. Bacterial biofilms: from the natural environment to infectious diseases // Nat. Rev. Microbiol. — 2004 Feb. — 2(2). — 95e108.
25. Hшiby N., Bjarnsholt T., Givskov M., Molin S., Ciofu O. Antibiotic resistance of bacterial biofilms // Int. J. Antimicrob Agents. — 2010. — 35. — 322e32.
26. Høiby N., Ciofu O., Johansen H.K., Song Z.J., Moser C., Jensen P.О. et al. The clinical impact of bacterial biofilms // Int. J. Oral Sci. — 2011. — 3. — 55e65.
27. Jakobsen T.H. et al. Qualitative and quantitative determination of quorum sensing inhibition in vitro. Quorum sensing: methods and protocols // Methods in Molecular Biology. — 2011. — Vol. 692. — P. 253-263.
28. Kostakioti M., Hadjifrangiskou M., Hultgren S.J. Bacterial biofilms: development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era // Cold Spring Harb. Perspect. Med. — 2013. — 3. — a010306.
29. Lambiase A., Rossano F., Piazza O., Del Pezzo M., Catania M.R., Tufano R. Typing of Pseudomonas aeruginosa isolated from patients with VAP in an intensive care unit // New Microbiol. — 2009. — 32. — 277e83.
30. Li F., Yu J., Yang H., Wan Z., Bai D. Effects of ambroxol on alginate of mature Pseudomonas aeruginosa biofilms // Curr. Microbiol. — 2008. — 57. — 1e7.
31. Lozano R., Naghavi M., Foreman K., Lim S., Shibuya K., Aboyans V. et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010 // Lancet. — 2012. — 380. — 2095e128.
32. Lu Q., Yu J., Bao L. et al. Effects of combined treatment with ambroxol and ciprofloxacin on catheter-associated Pseudomonas aeruginosa biofilms in a rat model // Chemotherapy. — 2013. — 59 (1). — 51-6; doi: 10.1159/000351107.
33. Lu Q., Yu J., Yang X., Wang J., Wang L., Lin Y. et al. Ambroxol interferes with Pseudomonas aeruginosa quorum sensing // Int. J. Antimicrob. Agents. — 2010. — 36. — 211e5.
34. Mah T.-F.C., O’Toole G.A. Mechanisms of biofilm resistance to antimicrobial agents // Trends Microbiol. — 2001. — 9. — 34e9.
35. Malerba M., Ragnoli B. Ambroxol in the 21st century: pharmacological and clinical update // Expert Opin. Drug Metab. Toxicol. — 2008. — 4. — 1119e29.
36. Mannino D.M. COPD: epidemiology, prevalence, morbidity and mortality, and disease heterogeneity // Chest. — 2002. — 121(Suppl. 5). — 121Se6S.
37. Masterton R.G., Galloway A., French G., Street M., Armstrong J., Brown E. et al. Guidelines for the management of hospital-acquired pneumonia in the UK: report of the working party on hospital-acquired pneumonia of the British Society for Antimicrobial Chemotherapy // J. Antimicrob. Chemother. — 2008. — 62. — 5e34.
38. Matthew R. Parsek Bacterial Biofilms: An Emerging Link to Disease Pathogenesis // Annual Review of Microbiology. — 2003. — Vol. 57. — 677-701.
39. McNabe M., Tennant R., Danelishvili L., Young L., Bermudez L.E. Mycobacterium avium ssp. hominissuis biofilm is composed of distinct phenotypes and influenced by the presence of antimicrobials // Clin. Microbiol. Infect. — 2011. — 17. — 697e703.
40. Mirsaeidi M., Hadid W., Ericsoussi B., Rodgers D., Sadikot R.T. Non-tuberculous mycobacterial disease is common in patients with non-cystic fibrosis bronchiectasis // Int. J. Infect. Dis. — 2013 Nov. — 17(11). — e1000e4.
41. Murphy T.F., Kirkham C., Sethi S., Lesse A.J. Expression of a peroxiredoxinglutaredoxin by Haemophilus influenzae in biofilms and during human respiratory tract infection // FEMS Immunol. Med. Microbiol. — 2005. — 44. — 81e9.
42. Muscedere J., Dodek P., Keenan S., Fowler R., Cook D., Heyland D. et al. Comprehensive evidence-based clinical practice guidelines for ventilatorassociated pneumonia: prevention // J. Crit. Care. — 2008. — 23. — 126e37.
43. Nazzari E., Torretta S., Pignataro L, Marchisio P., Esposito S. Role of biofilm in children with recurrent upper respiratory tract infections // European Journal of Clinical Microbiology & Infectious Diseases. — March 2015. — Vol. 34, Issue 3. — Р. 421-429.
44. Ndour C.T., Ahmed K., Nakagawa T., Nakano Y., Ichinose A., Tarhan G. et al. Modulating effects of mucoregulating drugs on the attachment of Haemophilus influenzae // Microb. Pathog. — 2001. — 30. — 121e7.
45. Nenke P. et al. The role of biofilm infection in urology // World Journal of Urology. — 2006. — Vol. 24. — Р. 13-20.
46. Nowak D., Antczak A., Krуl M. et al. Antioxidant properties of Ambroxol // Free Radic. Biol. Med. — 1994. — 16 (4). — 517-22.
47. O’Toole G., Kaplan H.B., Kolter R. Biofilm formation as microbial development // Annu Rev. Microbiol. — 2000. — 54. — 49e79.
48. Pintucci J.P., Corno S., Garotta M. Biofilms and infections of the upper respiratory tract // Eur. Rev. Med. Pharmacol. Sci. — 2010. — 14. — 683-90.
49. Pulcrano G., Panellis D., De Domenico G., Rossano F., Catania M.R. Ambroxol influences voriconazole resistance of Candida parapsilosis biofilm // FEMS Yeast Res. — 2012. — 12. — 430e8.
50. Ratjen F.A. Cystic fibrosis: pathogenesis and future treatment strategies // Respir. Care. — 2009. — 54. — 595e602.
51. Rene H.D., José M.S., Isela S.N., Claudio CR. Effects of ambroxol on Candida albicans growth and biofilm formation // Mycoses. — 2014 Apr. — 57(4). — 228-32.
52. Riise G.C., Qvarfordt I., Larsson S. et al. Inhibitory effect of N-acetylcysteine on adherence of Streptococcus pneumoniae and Haemophilus influenzae to human oropharyngeal epithelial cells in vitro // Respiration. — 2000. — 67. — 552e8.
53. Rovina N., Koutsoukou A., Koulouris N.G. Inflammation and immune response in COPD: where do we stand? // Mediators Inflamm. — 2013. — 2013. — 413735.
54. Saafan M.E., Ibrahim W.S., Tomoum M.O. Role of adenoid biofilm in chronic otitis media with effusion in children // Eur. Arch. Otorhinolaryngol. — 2013 Sep. — 270(9). — 2417-25. doi: 10.1007/s00405-012-2259-1. Epub 2012 Nov 21.
55. Torretta S., Drago L., Marchisio P., Cappadona M., Ri–naldi V., Nazzari Е., Pignataro L. Recurrences in chronic tonsillitis substained by tonsillar biofilm-producing bacteria in children // Relationship with the grade of tonsillar hyperplasy. — 2014. — Vol. 77, Issue 2. — P. 200-204.
56. Sauer K., Camper A.K., Ehrlich G.D. et al. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm // J. Bacteriol. — 2002. — 184 (4). — 1140-54.
57. Saylam G., Tatar E.C., Tatar I., Ozdek A., Korkmaz H. Association of adenoid surface biofilm formation and chronic otitis media with effusion // Arch. Otolaryngol. Head Neck Surg. — 2010 Jun. — 136 (6). — 550-5. doi: 10.1001/archoto.2010.70.
58. Schuster M., Sexton D.J., Diggle S.P., Greenberg E.P. Acyl-homoserine lactone quorum sensing: from evolution to application // Ann. Rev. Microbiol. — 2013. — 67. — 43e63.
59. Sethi S., Sethi R., Eschberger K., Lobbins P., Cai X., Grant B.J. et al. Airway bacterial concentrations and exacerbations of chronic obstructive pulmonary disease // Am. J. Respir. Crit. Care Med. — 2007. — 176. — 356e61.
60. Shan K.D. et al. Specialized persister cells and the mechanism of multidrug tolerance in Escherichia coli // J. Bacteriol. — 2004. — Vol. 186. — Р. 8172-8180.
61. Singh R. et al. Penetration of antibioties through Staphylococcus aureus and Staphylococcus epidermidis biofilms // J. Antimicrob. Chemother. — 2010. — Vol. 65. — Р. 1955-1958.
62. Snowden J.N., Beaver M., Smeltzer M.S., Kielian T. Biofilm-infected intracerebroventricular shunts elicit inflammation within the central nervous system // Infect. Immun. — 2012. — 80. — 3206e14.
63. Stewart P.S., Franklin M.J. Physiological hetero-geneity in biofilms // Nat. Rev. Microbiol. — 2008. — 6 (3). — 199-210.
64. Tetz V.V. et al. Extracellular phospholipids of isolated bacterial communities // Biofilms. — 2004. — Vol. 1. — Р. 149-155.
65. Tian X. Quorum sensing and bacterial social interactions in biofilms // Sensors (Basel). — 2012. — 12. — 2519e38.
66. Torres A., Ewig S., Lode H., Carlet J., European HAP working group. Defining, treating and preventing hospital acquired pneumonia: European perspective // Intensive Care Med. — 2009. — 35. — 9e29.
67. Varelogianni G., Hussain R., Strid H., Oliynyk I., Roomans G.M., Johannesson M. Effect of ambroxol on chloride transport, CFTR and ENaC in cystic fibrosis airway epithelial cells // Cell Biol. Int. — 2013 Jun. — 14. — http://dx.doi.org/10.1002/ cbin.
68. Vu B., Chen M., Crawford R.J. et al. Bacterial extracellular polysaccharides involved in biofilm formation // Molecules. — 2009. — 14. — 2535-54.
69. Vuong C., Voyich J.M., Fischer E.R., Braughton K.R., Whitney A.R., DeLeo F.R. et al. Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system // Cell Microbiol. — 2004. — 6. — 269e75.
70. Wiener-Kronish J.P., Dorr H.I. Ventilator-associated pneumonia: problems with diagnosis and therapy // Best Pract. Res Clin. Anaesthesiol. — 2008. — 22. — 437e49.
71. Yung-Hua Li. Xiaolin Tian. Quorum Sensing and Bacterial Social Interactions in Biofilms // Sensors. — 2012. — 12. — 2519-38.
72. Zhang B., Liu Y. Prophylaxis against ventilator-induced lung injury by Ambroxol // Zhonghua Yi Xue Za Zhi. — 2000. — 80 (1). — 51-3.
73. Zhang Y., Fu Y., Yu J., Ai Q., Li J., Peng N., Song S., He Y., Wang Z. Synergy of ambroxol with vancomycin in elimination of cathe–ter-related Staphylococcus epidermidis biofilm in vitro and in vivo // J. Infect Chemother. — 2015 Nov. — 21 (11). — 808-15.
74. Zhao S.P., Guo Q.L., Wang R.K., Wang E. Oxidative and anti-oxidative effects of ambroxol on acute hydrochloric acid-induced lung injury in rats // Zhong Nan Da Xue Xue Bao Yi Xue Ban. — 2004. — 29 (5). — 586-8.
75. Beeh К.М. et al. Anti- inflammatory properties of ambroxol // Eur. J. Med. Res. — 2008. — 13. — 557-562.
76. Банадига Н.В. Вибір препарату для лікування кашлю у дітей: доцільність, ефективність, безпечність, комплаєнс // Современная педиатрия. — 2015. — 1 (65). — 44-49.

/89.jpg)