
Журнал «Здоровье ребенка» Том 16, №1, 2021
Вернуться к номеру
Бактериальные поверхностно-активные вещества как агенты с антибиопленочной активностью
Авторы: Абатуров А.Е.
ГУ «Днепропетровская медицинская академия МЗ Украины», г. Днепр, Украина
Рубрики: Педиатрия/Неонатология
Разделы: Справочник специалиста
Версия для печати
Біосурфагенти являють собою гетерогенну групу біологічних поверхнево-активних амфіфільних сполук. Продуцентами біосурфагентів є різні мікроорганізми: бактерії і гриби. Клас біосурфагентів складається з двох груп: низькомолекулярних і високомолекулярних сполук. Представниками низькомолекулярних сполук є ліпопептиди, гліколіпіди, жирні кислоти, фосфоліпіди, що знижують поверхневий і міжфазний натяг, а високомолекулярних сполук — полімерні й дисперсні біосурфактанти, які являють собою емульсійні стабілізатори. Найбільш вивченими біосурфагентами, що мають потенціал лікарських засобів, є ліпопептиди і гліколіпіди. Підгрупу ліпопептидів представляють поліміксини, псевдофактини, путисолвіни, сурфактини, фенгіцин та інші, а гліколіпідів — рамноліпіди, трегалозні, софорозні, целобіозні, манозілеритритольні ліпіди та інші. Біосурфагенти відіграють ключову роль у життєдіяльності біоплівок: вони регулюють адгезію бактерій і матриксу біоплівки, підтримують функціонування каналів матриксу, забезпечуючи потреби бактерій у поживних речовинах. Також показано, що біосурфагенти беруть участь у формуванні та диспергуванні сформованих біоплівок. Дані речовини, прямо реагуючи з компонентами матриксу, індукують деградацію біоплівки. Біосурфагенти, маючи антимікробні, протигрибкові та противірусні властивості і поєднуючи антибактеріальну та антибіоплівкову дію, відкривають нові перспективи в терапії рецидивуючих, хронічних інфекційних захворювань. Вважають, що поверхнево-активні сполуки — як представники ліпопептидів, так і гліколіпідів — можуть бути молекулярною основою для розробки лікарських засобів, що будуть сприяти підвищенню ефективності антибактеріальної терапії проблемних інфекцій, особливо тих, що викликані антибіотико-резистентними штамами.
Биосурфагенты представляют собой гетерогенную группу биологических поверхностно-активных амфифильных соединений. Продуцентами биосурфагентов являются различные микроорганизмы: бактерии и грибы. Класс биосурфагентов состоит из двух групп: низкомолекулярных и высокомолекулярных соединений. Представителями низкомолекулярных соединений являются липопептиды, гликолипиды, жирные кислоты, фосфолипиды, которые снижают поверхностное и межфазное натяжение, а высокомолекулярных соединений — полимерные и дисперсные биосурфактанты, которые представляют собой эмульсионные стабилизаторы. Наиболее изученными биосурфагентами, обладающими потенциалом лекарственных средств, являются липопептиды и гликолипиды. Подгруппу липопептидов представляют полимиксины, псевдофактины, путисолвины, сурфактины, фенгицин и другие; а гликолипидов — рамнолипиды, трегалозные, софорозные, целлобиозные, маннозилеритритольные липиды и другие. Биосурфагенты играют ключевую роль в жизнедеятельности биопленок: они регулируют адгезию бактерий и матрикса биопленки, поддерживают функционирование каналов матрикса, обеспечивая потребности бактерий в питательных веществах. Также показано, что биосурфагенты участвуют в формировании и диспергировании сформированных биопленок. Данные вещества, прямо реагируя с компонентами матрикса, индуцируют деградацию биопленки. Биосурфагенты, обладая антимикробными, противогрибковыми и противовирусными свойствами и сочетанием антибактериального и антибиопленочного действия, открывают новые перспективы в терапии рецидивирующих, хронических инфекционных заболеваний. Полагают, что поверхностно-активные соединения — как представители липопептидов, так и гликолипидов — могут быть молекулярной основой для разработки лекарственных средств, которые будут способствовать повышению эффективности антибактериальной терапии проблемных инфекций, особенно тех, которые вызваны антибиотикорезистентными штаммами.
Biosuragents are a heterogeneous group of biological surface-active amphiphilic compounds. The producers of biosurfactants are various microorganisms: bacteria and fungi. The class of biosurfactants consists of two groups: low molecular weight and high molecular weight compounds. Representatives of low molecular weight compounds are lipopeptides, glycolipids, fatty acids, phospholipids that reduce surface and interfacial tension, and high molecular weight compounds are polymer and dispersed biosurfactants, which are emulsion stabilizers. The most studied biosurfactants with the potential of drugs are lipopeptides and glycolipids. A subgroup of lipopeptides are polymyxins, pseudo-factins, putisolvins, surfactin, fengycin and others; and glycolipids — rhamnolipids, trehalose, sophorose, cellobiose, mannosileritritol lipids, and others. Biosurfactants play a key role in the life of biofilms: they regulate the adhesion of bacteria and biofilm matrix, support the functioning of the matrix channels, providing the nutrient needs of bacteria. It has also been shown that biosurfactants are involved in the formation and dispersion of formed biofilms. These substances, directly reacting with the components of the matrix, induce degradation of the biofilm. Biosurfing agents, possessing antimicrobial, antifungal and antiviral, and antitumor properties, are a promising class of compounds that, possessing a combination of antibacterial and antibiofilm action, open up new perspectives in the treatment of recurrent chronic infectious diseases. It is believed that surface-active compounds, both representatives of lipopeptides and glycolipids, can be the molecular basis for the development of drugs that will enhance the effectiveness of antibiotic therapy for problem infections, especially those caused by antibiotic-resistant strains.
бактеріальні біоплівки; диспергування; біосурфагенти
бактериальные биопленки; диспергирование; биосурфагенты
bacterial biofilms; dispersion; biosurfagents
Введение
Микроорганизмы, продуцирующие биосурфагенты
/133.jpg)
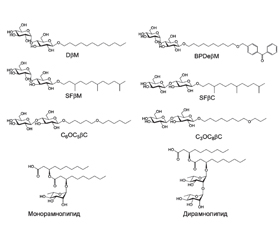
Липопептиды
/134.jpg)
/135.jpg)
/135_2.jpg)
/136.jpg)
/137.jpg)
/138.jpg)
/138_2.jpg)
/139.jpg)
Заключение
- Abdel-Mawgoud A.M., Lépine F., Déziel E. Rhamnolipids: diversity of structures, microbial origins and roles. Appl Microbiol. Biotechnol. 2010 May. 86 (5). 1323-36. doi: 10.1007/s00253-010-2498-2.
- Balleza D., Alessandrini A., Beltrán García M.J. Role of Lipid Composition, Physicochemical Interactions, and Membrane Mechanics in the Molecular Actions of Microbial Cyclic Lipopeptides. J. Membr. Biol. 2019 Jun. 252 (2–3). 131-157. doi: 10.1007/s00232-019-00067-4.
- Banat I.M., De Rienzo M.A., Quinn G.A. Microbial biofilms: biosurfactants as antibiofilm agents. Appl Microbiol. Biotechnol. 2014 Dec. 98 (24). 9915-29. doi: 10.1007/s00253-014-6169-6.
- Banat I.M., Franzetti A., Gandolfi I., Bestetti G., Martinotti M.G., Fracchia L., Smyth T.J., Marchant R. Microbial biosurfactants production, applications and future potential. Appl Microbiol. Biotechnol. 2010 Jun. 87 (2). 427-44. doi: 10.1007/s00253-010-2589-0.
- Berditsch M., Jäger T., Strempel N., Schwartz T., Overhage J., Ulrich A.S. Synergistic effect of membrane-active peptides polymyxin B and gramicidin S on multidrug-resistant strains and biofilms of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015 Sep. 59 (9). 5288-96. doi: 10.1128/AAC.00682-15.
- Bhattacharjee A., Nusca T.D., Hochbaum A.I. Rhamnolipids Mediate an Interspecies Biofilm Dispersal Signaling Pathway. ACS Chem. Biol. 2016 Nov. 18. 11 (11). 3068-3076. doi: 10.1021/acschembio.6b00750.
- Biniarz P., Coutte F., Gancel F., Łukaszewicz M. High-throughput optimization of medium components and culture conditions for the efficient production of a lipopeptide pseudofactin by Pseudomonas fluorescens BD5. Microb. Cell. Fact. 2018 Aug. 4. 17 (1). 121. doi: 10.1186/s12934-018-0968-x.
- Bionda N., Fleeman R.M., de la Fuente-Núñez C., Rodriguez M.C., Reffuveille F., Shaw L.N., Pastar I., Davis S.C., Hancock R.E.W., Cudic P. Identification of novel cyclic lipopeptides from a positional scanning combinatorial library with enhanced antibacterial and antibiofilm activities. Eur. J. Med. Chem. 2016 Jan. 27. 108. 354-363. doi: 10.1016/j.ejmech.2015.11.032.
- Brasseur R., Braun N., El Kirat K., Deleu M., Mingeot-Leclercq M.P., Dufrêne Y.F. The biologically important surfactin lipopeptide induces nanoripples in supported lipid bilayers. Langmuir. 2007 Sep. 11. 23 (19). 9769-72. doi: 10.1021/la7014868.
- Ceresa C., Tessarolo F., Maniglio D., Tambone E., Carmagnola I., Fedeli E., Caola I., Nollo G., Chiono V., Allegrone G., Rinaldi M., Fracchia L. Medical-Grade Silicone Coated with Rhamnolipid R89 Is Effective against Staphylococcus spp. Biofilms. Molecules. 2019 Oct. 25. 24 (21). pii: E3843. doi: 10.3390/molecules24213843.
- Díaz De Rienzo M.A., Banat I.M., Dolman B., Winterburn J., Martin P.J. Sophorolipid biosurfactants: Possible uses as antibacterial and antibiofilm agent. N. Biotechnol. 2015 Dec. 25. 32 (6). 720-6. doi: 10.1016/j.nbt.2015.02.009.
- Diaz De Rienzo M.A., Stevenson P.S., Marchant R., Banat I.M. Effect of biosurfactants on Pseudomonas aeruginosa and Staphylococcus aureus biofilms in a BioFlux channel. Appl Microbiol. Biotechnol. 2016 Jul. 100 (13). 5773-9. doi: 10.1007/s00253-016-7310-5.
- Ezadi F., Ardebili A., Mirnejad R. Antimicrobial Susceptibility Testing for Polymyxins: Challenges, Issues, and Recommendations. J. Clin. Microbiol. 2019 Mar. 28. 57 (4). pii: e01390-18. doi: 10.1128/JCM.01390-18.
- Fariq A., Saeed A. Production and Biomedical Applications of Probiotic Biosurfactants. Curr. Microbiol. 2016 Apr. 72 (4). 489-95. doi: 10.1007/s00284-015-0978-4.
- Fenibo E.O., Ijoma G.N., Selvarajan R., Chikere C.B. Microbial Surfactants: The Next Generation Multifunctional Biomolecules for Applications in the Petroleum Industry and Its Associated Environmental Remediation. Microorganisms. 2019 Nov. 19. 7 (11). pii: E581. doi: 10.3390/microorganisms7110581.
- Fernández-Barat L., Ciofu O., Kragh K.N., Pressler T., Johansen U., Motos A., Torres A., Hoiby N. Phenotypic shift in Pseudomonas aeruginosa populations from cystic fibrosis lungs after 2-week antipseudomonal treatment. J. Cyst. Fibros. 2017 Mar. 16 (2). 222-229. doi: 10.1016/j.jcf.2016.08.005.
- Gudiña E.J., Rangarajan V., Sen R., Rodrigues L.R. Potential therapeutic applications of biosurfactants. Trends Pharmacol. Sci. 2013 Dec. 34 (12). 667-75. doi: 10.1016/j.tips.2013.10.002.
- Hajfarajollah H., Eslami P., Mokhtarani B., Akbari Noghabi K. Biosurfactants from probiotic bacteria: A review. Biotechnol. Appl Biochem. 2018 Nov. 65 (6). 768-783. doi: 10.1002/bab.1686.
- Hanif A., Zhang F., Li P., Li C., Xu Y., Zubair M., Zhang M., Jia D., Zhao X., Liang J., Majid T., Yan J., Farzand A., Wu H., Gu Q., Gao X. Fengycin Produced by Bacillus amyloliquefaciens FZB42 Inhibits Fusarium graminearum Growth and Mycotoxins Biosynthesis. Toxins (Basel). 2019 May 24. 11 (5). pii: E295. doi: 10.3390/toxins11050295.
- He J., Ledesma K.R., Lam W.Y., Figueroa D.A., Lim T.P., Chow D.S., Tam V.H. Variability of polymyxin B major components in commercial formulations. Int. J. Antimicrob. Agents. 2010 Mar. 35 (3). 308-10. doi: 10.1016/j.ijantimicag.2009.11.005.
- Inès M., Dhouha G. Glycolipid biosurfactants: Potential related biomedical and biotechnological applications. Carbohydr. Res. 2015 Oct. 30. 416. 59-69. doi: 10.1016/j.carres.2015.07.016.
- Inès M., Dhouha G. Lipopeptide surfactants: Production, recovery and pore forming capacity. Peptides. 2015 Sep. 71. 100-12. doi: 10.1016/j.peptides.2015.07.006.
- Irie Y., O'toole G.A., Yuk M.H. Pseudomonas aeruginosa rhamnolipids disperse Bordetella bronchiseptica biofilms. FEMS Microbiol Lett. 2005 Sep. 15. 250 (2). 237-43. doi: 10.1016/j.femsle.2005.07.012.
- Janek T., Lukaszewicz M., Krasowska A. Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbiol. 2012 Feb. 23. 12. 24. doi: 10.1186/1471-2180-12-24.
- Janek T., Lukaszewicz M., Rezanka T., Krasowska A. Isolation and characterization of two new lipopeptide biosurfactants produced by Pseudomonas fluorescens BD5 isolated from water from the Arctic Archipelago of Svalbard. Bioresour. Technol. 2010 Aug. 101 (15). 6118-23. doi: 10.1016/j.biortech.2010.02.109.
- Jezierska S., Claus S., Van Bogaert I. Yeast glycolipid biosurfactants. FEBS Lett. 2018 Apr. 592 (8). 1312-1329. doi: 10.1002/1873-3468.12888.
- Klinger-Strobel M., Stein C., Forstner C., Makarewicz O., Pletz M.W. Effects of colistin on biofilm matrices of Escherichia coli and Staphylococcus aureus. Int. J. Antimicrob. Agents. 2017 Apr. 49 (4). 472-479. doi: 10.1016/j.ijantimicag.2017.01.005.
- Kolpen M., Appeldorff C.F., Brandt S., Mousavi N., Kragh K.N., Aydogan S., Uppal H.A., Bjarnsholt T., Ciofu O., Hoiby N., Jensen P.O. Increased bactericidal activity of colistin on Pseudomonas aeruginosa biofilms in anaerobic conditions. Pathog. Dis. 2016 Feb. 74 (1). ftv086. doi: 10.1093/femspd/ftv086.
- Krasowska A., Lukaszewicz M., Grzywacz D., Kamysz W. A novel method of pseudofactin synthesis https//patents.google.com/patent/EP3000822A1/en.
- Kulakovskaya E., Kulakovskaya T. Structure and Occurrence of Yeast Extracellular Glycolipids. Extracellular Glycolipids of Yeasts. 2014.
- Lora-Tamayo J., Murillo O., Ariza J. Clinical Use of Colistin in Biofilm-Associated Infections. Adv. Exp. Med. Biol. 2019. 1145. 181-195. doi: 10.1007/978-3-030-16373-0_13.
- Malinowski A.M., McClarty B.M., Robinson C., Spear W., Sanchez M., Sparkes T.C., Brooke J.S. Polysorbate 80 and polymyxin B inhibit Stenotrophomonas maltophilia biofilm. Diagn. Microbiol. Infect. Dis. 2017 Feb. 87 (2). 154-156. doi: 10.1016/j.diagmicrobio.2016.11.008.
- Michalopoulos A., Falagas M.E. Colistin and polymyxin B in critical care. Crit. Care Clin. 2008 Apr. 24 (2). 377-91, x. doi: 10.1016/j.ccc.2007.12.003.
- Mnif I., Ghribi D. Glycolipid biosurfactants: main properties and potential applications in agriculture and food industry. J. Sci Food Agric. 2016 Oct. 96 (13). 4310-20. doi: 10.1002/jsfa.7759.
- Mongkolthanaruk W. Classification of Bacillus beneficial substances related to plants, humans and animals. J. Microbiol. Biotechnol. 2012 Dec. 22 (12). 1597-604. doi: 10.4014/jmb.1204.04013.
- Moradali M.F., Ghods S., Rehm B.H. Pseudomonas aeruginosa Lifestyle: A Paradigm for Adaptation, Survival, and Persistence. Front Cell Infect. Microbiol. 2017 Feb. 15. 7. 39. doi: 10.3389/fcimb.2017.00039.
- Ongena M., Jourdan E., Adam A., Paquot M., Brans A., Joris B., Arpigny J.L., Thonart P. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ Microbiol. 2007 Apr. 9 (4). 1084-90. doi: 10.1111/j.1462-2920.2006.01202.x.
- Oppenheimer-Shaanan Y., Steinberg N., Kolodkin-Gal I. Small molecules are natural triggers for the disassembly of biofilms. Trends Microbiol. 2013 Nov. 21 (11). 594-601. doi: 10.1016/j.tim.2013.08.005.
- Otzen D.E. Biosurfactants and surfactants interacting with membranes and proteins: Same but different? Biochim. Biophys. Acta Biomembr. 2017 Apr. 1859 (4). 639-649. doi: 10.1016/j.bbamem.2016.09.024.
- Poirel L., Jayol A., Nordmann P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017 Apr. 30 (2). 557-596. doi: 10.1128/CMR.00064-16.
- Raaijmakers J.M., De Bruijn I., Nybroe O., Ongena M. Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol. Rev. 2010 Nov. 34 (6). 1037-62. doi: 10.1111/j.1574-6976.2010.00221.x.
- Rivardo F., Turner R.J., Allegrone G., Ceri H., Martinotti M.G. Anti-adhesion activity of two biosurfactants produced by Bacillus spp. prevents biofilm formation of human bacterial pathogens. Appl Microbiol. Biotechnol. 2009 Jun. 83 (3). 541-53. doi: 10.1007/s00253-009-1987-7.
- Ron E.Z., Rosenberg E. Natural roles of biosurfactants. Environ Microbiol. 2001 Apr. 3 (4). 229-36. doi: 10.1046/j.1462-2920.2001.00190.x.
- Roy A., Mahata D., Paul D., Korpole S., Franco O.L., Mandal S.M. Purification, biochemical characterization and self-assembled structure of a fengycin-like antifungal peptide from Bacillus thuringiensis strain SM1. Front Microbiol. 2013 Nov. 21. 4. 332. doi: 10.3389/fmicb.2013.00332.
- Salman M., Rizwana R., Khan H., Munir I., Hamayun M., Iqbal A., Rehman A., Amin K., Ahmed G., Khan M., Khan A., Amin F.U. Synergistic effect of silver nanoparticles and polymyxin B against biofilm produced by Pseudomonas aeruginosa isolates of pus samples in vitro. Artif. Cells Nanomed. Biotechnol. 2019 Dec. 47 (1). 2465-2472. doi: 10.1080/21691401.2019.1626864.
- Santos D.K., Rufino R.D., Luna J.M., Santos V.A., Sarubbo L.A. Biosurfactants: Multifunctional Biomolecules of the 21st Century. Int. J. Mol. Sci. 2016 Mar. 18. 17 (3). 401. doi: 10.3390/ijms17030401.
- Satputea S.K., Banpurkar A.G., Banat I.M. et al. Multiple Roles of Biosurfactants in Biofilms. Curr. Pharm. Des. 2016. 22 (11). 1429-48. doi: 10.2174/1381612822666160120152704.
- Schneider-Futschik E.K., Paulin O.K.A., Hoyer D., Roberts K.D., Ziogas J., Baker M.A., Karas J., Li J., Velkov T. Sputum Active Polymyxin Lipopeptides: Activity against Cystic Fibrosis Pseudomonas aeruginosa Isolates and Their Interactions with Sputum Biomolecules. ACS Infect. Dis. 2018 May 11. 4 (5). 646-655. doi: 10.1021/acsinfecdis.7b00238.
- Vanittanakom N., Loeffler W., Koch U., Jung G. Fengycin — a novel antifungal lipopeptide antibiotic produced by Bacillus subtilis F-29-3. J. Antibiot. (Tokyo). 1986 Jul. 39 (7). 888-901. doi: 10.7164/antibiotics.39.888.
- Vollenbroich D., Ozel M., Vater J., Kamp R.M., Pauli G. Mechanism of inactivation of enveloped viruses by the biosurfactant surfactin from Bacillus subtilis. Biologicals. 1997 Sep. 25 (3). 289-97. doi: 10.1006/biol.1997.0099.
- Vuotto C., Donelli G. Novel Treatment Strategies for Biofilm-Based Infections. Drugs. 2019 Oct. 79 (15). 1635-1655. doi: 10.1007/s40265-019-01184-z.
- Wang J., Yu B., Tian D., Ni M. Rhamnolipid but not motility is associated with the initiation of biofilm seeding dispersal of Pseudomonas aeruginosa strain PA17. J. Biosci. 2013 Mar. 38 (1). 149-56. doi: 10.1007/s12038-012-9297-0.
- Wood T.L., Gong T., Zhu L., Miller J., Miller D.S., Yin B., Wood T.K. Rhamnolipids from Pseudomonas aeruginosa disperse the biofilms of sulfate-reducing bacteria. NPJ Biofilms Microbiomes. 2018 Oct. 3. 4. 22. doi: 10.1038/s41522-018-0066-1.
- Ye L., Hildebrand F., Dingemans J., Ballet S., Laus G., Matthijs S., Berendsen R., Cornelis P. Draft genome sequence analysis of a Pseudomonas putida W15Oct28 strain with antagonistic activity to Gram-positive and Pseudomonas sp. pathogens. PLoS One. 2014 Nov. 4. 9 (11). e110038. doi: 10.1371/journal.pone.0110038.
- Zemenová J., Sýkora D., Maletínská L., Kuneš J. Lipopeptides as therapeutics: applications and in vivo quantitative analysis. Bioanalysis. 2017 Jan. 9 (2). 215-230. PMID: 27960546.
- Zhao H., Shao D., Jiang C., Shi J., Li Q., Huang Q., Rajoka M.S.R., Yang H., Jin M. Biological activity of lipopeptides from Bacillus. Appl Microbiol. Biotechnol. 2017 Aug. 101 (15). 5951-5960. doi: 10.1007/s00253-017-8396-0.
- Zheng H., Singh N., Shetye G.S. et al. Synthetic analogs of rhamnolipids modulate structured biofilms formed by rhamnolipid-nonproducing mutant of Pseudomonas aeruginosa. Bioorg Med. Chem. 2017 Mar. 15. 25 (6). 1830-1838. doi: 10.1016/j.bmc.2017.01.042.